Document Actions
Live-cell imaging and analysis
In recent years there has been a revolution in analysing the cell biology of living cells. This has been mainly due to the development of: a wide range of fluorescent probes which can be used to non-invasively interrogate living cells; new microscope technologies (e.g. confocal microscopy, multiphoton microscopy, and fluorescence lifetime imaging microscopy [FLIM]); and computer software and hardware for digital image processing and analysis. These innovations are having a profound impact on the experimental analysis of living cells and are providing exciting and novel insights into their cell biology. Analysing the cell biology of living cells involves a fundamentally different philosophical and experimental approach to that involving the biochemical analysis of fractionated cell extracts from bulk multicellular systems or using microscopical methods which require the fixation of cells.
Important advantages of studying living cells include:
- analysing living cells produces more direct and therefore more reliable data for understanding living processes;
- these processes can be analysed in the same cells continuously or at defined intervals rather than periodically sampling different cells or cell populations;
- a large range of fluorescent and luminescent probes offer unparalleled opportunities for visualizing and quantifying the temporal and spatial dynamics of molecules, organelles, and physiological activities within living cells.
A major advantage of the analysis of single living cell is that it can reveal variation within a cell population which is missed when average measurements from across a whole cell population are obtained (as arises when using cell extracts from bulk samples).
A primary aim of our work in COSMIC is to push live-cell imaging to its limits with laser scanning confocal microscopy, Nipkov disc confocal microscopy, multiphoton microscopy, and FLIM. An important feature of the interdisciplinary COSMIC facility is that it includes Category 2 laboratories for the culturing of fungal, plant and animal cells.
Projects in COSMIC will include:
- Protein-protein interactions and secretory vesicle trafficking - We are addressing the dynamics of protein-protein interactions and secretory vesicle trafficking in mammalian cells through the integration of molecular approaches with high resolution cellular imaging techniques. Current projects include: (a) Regulation of protein-protein interactions involved in regulated exocytosis; (b) Analysis of single vesicle dynamics in living cells; (c) Colocalisation and protein-interaction studies of ion channels. Using a combination of confocal microscopy, multiphoton microscopy and FLIM we are addressing the in vivo real-time dynamics of regulated protein-protein interactions involved in the regulation of ion channels as well as regulated exocytosis in living cells. This allows us to monitor the three dimensional distribution, colocalisation and/or interaction of protein partners in real time
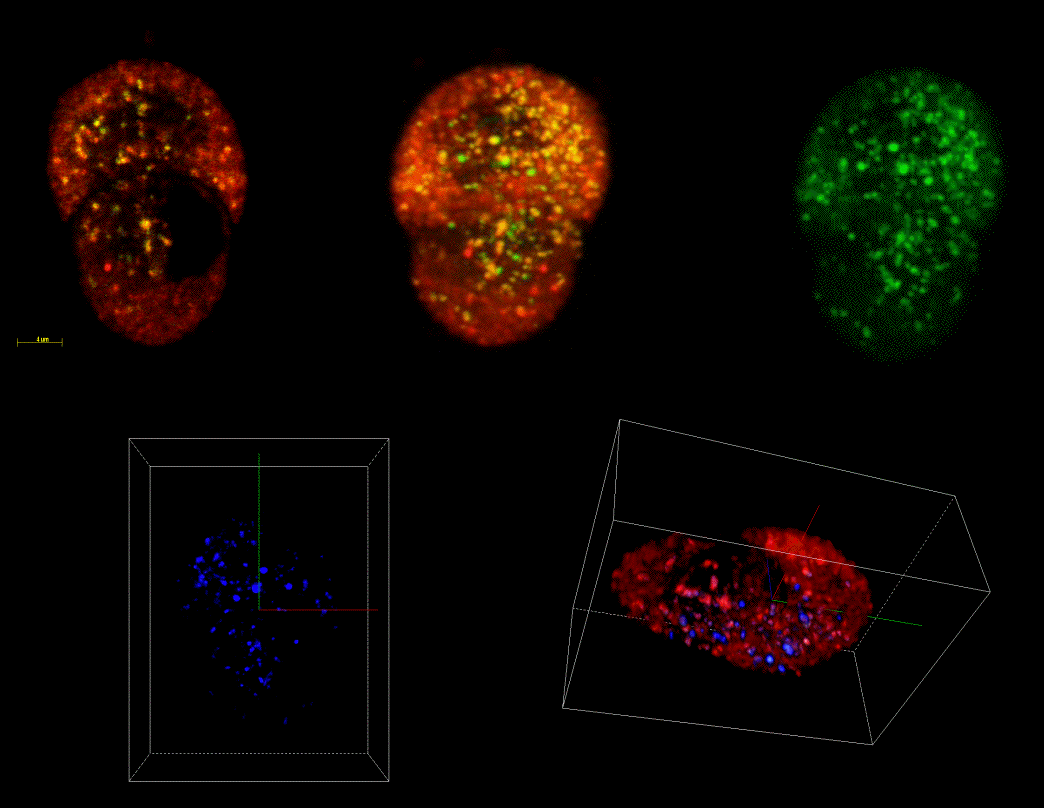
Fig 1 3-Dimensional confocal reconstruction of secretory vesicle marker colocalisation in a living endocrine cell. Secretory vesicles were targeted with an EGFP-ANF fusion protein (green) and acidic compartments labelled with lysotracker. Colocalisation is observed towards the base of the cell (blue) in structures with size distribution similar to secretory vesicles - click the image for a larger version.
In addition, targeted GFP-fusion protein are being used to specifically label secretory vesicles in neuroendocrine cells to monitor the real time dynamics of vesicle mobility and fusion. This allows us to label a limited subset of vesicles whose cellular distribution and mobility can be tracked (Fig. 2) and, with the use of total internal reflection microscopy (TIRFM), allows us to monitor the final fusion event in real time (Fig. 3).
- Live-cell imaging of hyphal fusion in a filamentous fungus (P.C.Hickey and N.D. Read) - Colony morphogenesis in filamentous fungi consists of three integrated processes: hyphal tip extension, branching, and fusion. Of these, hyphal fusion is the least understood. We have studied the process of hyphal fusion (anastomosis) in growing colonies of Neurospora crassa, stained with the membrane-selective dyes FM1-43 and FM4-64, and visualized by confocal microscopy using the BioRad laser scanning confocal microscope facility in COSMIC. Time-lapse, live-cell imaging illustrated the dynamics of hyphal growth and a nastomosis during its pre-contact, contact and post-contact, and post-fusion stages. Fusion-competent hyphae were morphologically distinct and exhibited remote sensing, resulting in branch initiation and/or re-direction of growth to facilitate contact between participating hyphae (Fig. 2). A stained "Spitzenkörper" was often observed where fusion-competent hyphae met. Our results provide evidence that this structure contains secretory vesicles responsible for the delivery of cell adhesion molecules at the point of contact, cell wall synthesizing enzymes for the swelling growth of fused hyphal tips, and digestive enzymes required for fusion pore formation. Dramatic changes in cytoplasmic flow frequently occurred between the participating hyphae following fusion. After anastomosis has taken place, septa commonly formed close to the fusion site. The live-cell imaging which we have reported has clearly shown the complexity of the hyphal homing and fusion process. The research conducted here was part of a collaboration with Prof. Louise Glass's group at the University of California Berkeley and has been recently published (Hickey et al., 2002). It was partly funded by the agrochemical company Syngenta.
- Hyphal tip growth in fungi - A defining feature of the fungal kingdom is the predominance of tip growth, a process which is responsible for most of the fungal biomass on this planet. Tip growth and the way it is regulated, often in quite subtle ways, is of critical importance for fungal morphogenesis, including the growth of filamentous cells (hyphae) and yeast cells, and the differentiation of spores and infection structures. The fungal lifestyle is thus crucially dependent on the pattern of tip growth. Vesicle trafficking underpins this process. Although the significance of polarized secretion has been long appeciated as central to tip growth, only recently has it emerged that endocytosis also plays major roles in it. Vesicles and organelles within fungal hyphae can be readily labelled with fluorescent dyes
(fig. 4 - click on the image to play movie)
or GFP
(fig. 5 - click on the image to play movie)
Confocal microscopy, multiphoton microscopy, Nipkov disc confocal microscopy and FLIM will be used to image the vesicle trafficking network in wild type and mutant strains of a variety of fungi, including plant and human pathogens
(fig. 6 - click on the image to play movie)
These studies will be combined with the manipulation of the apical cluster of secretory vesicles (Fig. 4) using laser tweezers in order to experimentally perturb the pattern of tip growth and branch formation.
- Touch-sensing by fungi - Some plant pathogens (e.g. rust fungi) have evolved a highly sophisticated mechanism for sensing microtopographical signals on a leaf surface in order to direct the growth of their hyphae towards sites of infection (stomata). Our previous work has shown that the microtopographical signals on a leaf can be mimicked on microfabricated substrata, and can be used to precisely regulate the direction of hyphal growth, sites of branch formation and the induction of infection structures over stomata. Using substrates with precisely microfabricated topographies and imprinting defined patterns of surface properties (e.g. surface chemistry and hydrophobicity) should allow predetermined hyphal network patterns to be generated. These studies will be extended to other genetically more amenable fungi, and combined with live-cell imaging using confocal microscopy, multiphoton microscopy and FLIM in order to understand the mechanistic basis of the signalling processes involved.
- Calcium signalling - Calcium plays a highly versatile and pivotal role as an intracellular signalling molecule, and is able to transduce a wide range of external chemical and physical signals through different signalling pathways, resulting in a variety of cellular responses. In COSMIC, confocal microscopy, multiphoton microscopy, Nipkov disc confocal microscopy, FLIM and luminescence imaging will be used to image, analyse and understand better the spatial characteristics of calcium during plant and fungal cell signalling and the downstream processes that interpret calcium signals. Calcium-sensitive fluorescent probes and recombinant aequorin will be used for this purpose. Our belief and long term goal is to establish that spatial calcium characteristics represent a kind of intracellular neural network capable of computation and integration of information from a multiplicity of signals impinging on a plant or fungal cell that enable apparent teleological and indeed rational responses to external signals.
Sources of funding: Wellcome; BBSRC; Syngenta, NATO, MAFF.
Fungal Cell Biology Group website: http://www.fungalcell.com
Membrane Biology Group website: http://www.bms.ed.ac.uk/mbg/index.htm
Molecular Signalling Group website: https://www.ed.ac.uk/~gidi/main.html
Collaborators: University of California at Berkeley, Cornell University, University of Bangor, University of York, University of Lisbon.
Fig 1: 3-Dimensional confocal reconstruction of secretory vesicle marker colocalisation in a living endocrine cell. Secretory vesicles were targeted with an EGFP-ANF fusion protein (green) and acidic compartments labelled with lysotracker. Colocalisation is observed towards the base of the cell (blue) in structures with size distribution similar to secretory vesicles.
Fig. 2: Tracking of single secretory vesicels in a living endocrine cell. Confocal 3D-reconstructions of a limited number (~150) of secretory vesicles labelled with EGFP-ANF before and after stimulation of secretion from the cell. Note the loss of vesicles after stimulation.
Fig. 3. Reconstruction of a single secretory vesicle fusion event in a living endocrine cell. Serial time-course images of a single secretory vesicle in an endocrine cell were captured using TIRFM and images stacked, and surface rendered to give a 2D+1 image reconstruction with time in the z-axis. The stalk represents an immobile vesicle before fusion to the plasma membrane, the ‘mushroom cap’ results from vesicle fusion and release and radial spread of its contents (EGFP-ANF) into the extracellular space towards the evanescent wave.
Fig. 4. Growing hyphal tip of the plant pathogen Sclerotinia sclerotiorum stained with FM4-64 and imaged by confocal microscopy. Note the strongly stained cluster of secretory vesicles in the hyphal tip.
Fig. 5. Dividing nuclei in hyphae of Aspergillus nidulans expressing nuclear-targeted GFP and imaged by confocal microscopy.
Fig. 6. Asexual structures of the human pathogen Aspergillus fumigatus imaged by confocal microscopy. Movie shows optical sectioning and 3D reconstruction.